按关键词阅读:
在病毒感染横行霸道的今天 , 免疫记忆是机体为防止感染的反复发生所必需的环节 , 如今所有的有效疫苗都是在此基础上展开研制 , 并在接种后对人体起到显著效果的 。 最有效的疫苗会引发记忆B细胞在病原体攻击后产生快速的记忆性体液免疫反应 。 然而 , 并不是所有疫苗都同样有效或能产生持续的中和反应 , 比如HIV-1疫苗的研发障碍 , 在某种程度上是因为人们几乎不了解表达中和抗体的记忆B细胞是如何为响应一系列相关病毒或序列免疫原而选择和进化的【1】【2】【3】 。
于是 , Charlotte Viant等人对此展开了研究 , 并于近日在《Cell》上发表了最新成果:Antibody Affinity Shapes the Choice between Memory and Germinal Center B Cell Fates 。 结合谱系溯源和单克隆B细胞的抗体来研究亲和性在B细胞选择生发中心(GCs)和免疫HIV-1抗原小鼠的记忆B细胞室中的作用 。

文章图片
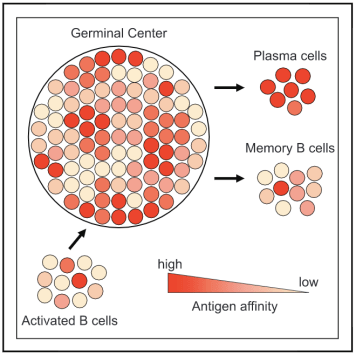
实验中 , Viant等人在具有完整免疫系统的免疫小鼠中 , 采用了无偏命运映射的方法 。 用HIV-1抗原对小鼠进行免疫 , 以了解B细胞进入记忆区室和同期的生发中心(GCs)B细胞之间的关系 。 实验结果表明 , GC中的大部分B细胞表达高亲和力的B细胞受体 , 这些受体源自具有显著亲和力的UCAs;相反 , 大多数同期选择的记忆B细胞则表达对抗原亲和力极低的受体 。
文章图片
单细胞转移试验表明 , 在细胞分裂后 , 单个前体会引起包括记忆B细胞在内的多个B细胞命运 。 通过抗原结合进行鉴定时 , 产生浆细胞和记忆细胞的前体比只产生浆细胞的前体结合的抗原数量更少 。 此外 , 转基因B细胞在受到较低亲和力抗原的攻击时 , 会产生更多的记忆B细胞【4】 。
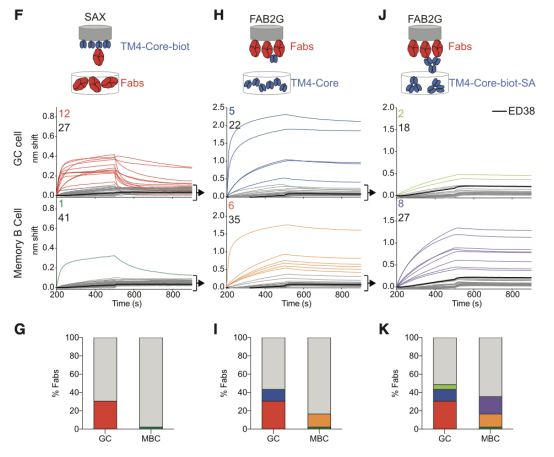
Viant等人则直接比较了同期的记忆和GC区室B细胞表达的抗体的亲和力 。 在单价结合条件下 , 有30.7%的GC和仅2.3%的记忆B细胞显示出可测结合;而在具有人工增加的化合价的测定中 , 将Fabs固定在传感芯片上 , 并暴露于三聚体抗原的四聚体中 , 这些数量分别增加至48.7%和35.7% 。 由此得出 , 新生的记忆细胞与抗原结合 , 但需要高价相互作用 , 通过亲和力效应增加表观亲和力 , 导致区室的整体亲和力远低于同期GC 。
然而 , 这种情况不足以代表体内B细胞表面高密度二价受体与免疫复合物相互作用的情况 , 因此无法检测在高价态的低亲和力相互作用 。 此外 , GC和记忆B细胞之间的差异与体细胞突变的数量无关 , 因为在比较5个或更少突变的GC和记忆B细胞时 , 亲和力的差异也可见 。
文章图片
绝大多数的记忆B细胞区室由表达低亲和力受体的细胞组成 , 但在人和小鼠的记忆区室中也包含少量携带高亲和力抗体的B细胞 , 这些抗体可以通过其结合能力来鉴定抗原 。 在感染HIV-1的人类中 , 记忆B细胞就表现出高度的抗原结合和中和活性【5】 。 事实上 , Viant等人的发现与早期实验并不矛盾 , 关键区别在于命运映射方法在结合抗原的记忆B细胞的选择时无偏 。
实验观察到 , GC中最大的扩展克隆在记忆区室中并不具有代表性 。 在GC中 , 细胞基于结合、内化和呈现抗原的能力竞争以限制T卵泡辅助细胞(Tfh)的帮助 。 与Tfh细胞的相互作用导致基因表达程序的激活 , 该程序取决于抗原呈现的数量 。 而接受更高水平帮助的B细胞上调增殖所需的基因或浆细胞分化 , 较低水平的抗原呈现导致更少的分裂 , 并减少与记忆B细胞发育相关的Bach2表达 。
此外 , 实验数据表明 , 在免疫反应的后期阶段 , 进入记忆区室的具有较高突变数量的B细胞几乎没有甚至只有一点 , 这一现象不会因反复免疫而改变 。 由于突变与细胞分裂直接相关 , GC细胞每4-6小时分裂一次 , 因此突变数量相对较低 , 这表明在免疫反应的后期阶段 , 大多数加入记忆区室的B细胞在GC中分裂所花费的时间很少 。 ![]()
稿源:(健康界)
【】网址:http://www.shadafang.com/c/hn1102c19252020.html
标题:Cell|Cell重磅 | 记忆和生发中心B细胞的命运选择由抗体亲和力决定!
